
iNature
截止2019年10月25日,浙江大学在Cell,Nature及Science上发表了9篇重要研究成果,iNature系统总结了这些成果:
【1】阳离子-氯化物共转运蛋白(CCC)介导阳离子与氯化物在质膜上的耦合,电中性共价传递,对于细胞体积调节,肾脏盐分重吸收以及γ-氨基丁酸(GABA)介导的神经元调节至关重要。 2019年10月25日,浙江大学郭江涛,李敬源,天津大学叶升,德克萨斯大学西南医学中心白晓辰及范德比尔特大学的Eric Delpire共同通讯在Science 发表题为“Cryo-EM structures of the human cation-chloride cotransporter KCC1”的研究论文,该研究介绍了在2.9至3.5埃分辨率下,在氯化钾或氯化钠中人氯化钾共转运蛋白KCC1的冷冻电子显微镜(cryo-EM)结构。KCC1以二聚体形式存在,胞外和跨膜结构域均参与二聚化。结构和功能分析以及计算研究表明,KCC1中有一个钾位和两个氯化物位,这些都是离子迁移活性所必需的。KCC1采用向内构象,并封闭了细胞外门。KCC1结构使我们能够对KCC中潜在的离子传输机制进行建模,并为药物设计提供蓝图;【2】核苷酸寡聚化域(NOD)样受体1和2(NOD1 / 2)是细胞内模式识别蛋白,可响应与微生物相关的肽聚糖而激活免疫信号通路。NOD1 / 2信号传导需要募集含细菌的内体和其他细胞内膜,而破坏膜定位的NOD1 / 2突变与炎症性肠病和其他炎症性疾病有关。但是,对于这一招募过程知之甚少。2019年10月25日,浙江大学医学院基础医学Dante Neculai、孙启明、加拿大Princess Margaret癌症研究所研究中心Brain Raught及多伦多St Michael医院Gregory D. Fairn共同通讯在Science在线发表题为“Palmitoylation of NOD1 and NOD2 is required for bacterial sensing”的研究论文,该研究发现膜募集和免疫信号传导需要NOD1 / 2 S-棕榈酰化。ZDHHC5被确定为负责此关键的翻译后修饰的棕榈酰转移酶,并且发现NOD2中的几种与疾病相关的突变与缺陷性S-棕榈酰化有关。因此,ZDHHC5介导的NOD1 / 2的S-棕榈酰化对于它们对肽聚糖的应答和进行有效免疫应答的能力至关重要。
【3】高熵合金是一类材料,其中包含五个或更多近似等原子比例的元素。它们非常规的成分和化学结构有望实现前所未有的机械性能组合。这类合金的合理设计取决于对几乎无限的组成空间中的组成,结构和性能关系的理解。2019年10月9日,浙江大学电子显微镜中心余倩,乔治亚理工Ting Zhu、加州大学伯克利分校Robert Ritchie共同通讯在Nature在线发表题为"Tuning element distribution, structure and properties by composition in high-entropy alloys"的研究论文,该研究使用原子分辨率化学映射来揭示广泛研究的面心立方CrMnFeCoNi Cantor合金和新面心立方合金CrFeCoNiPd的元素分布。在Cantor合金中,五个组成元素的分布相对随机且均匀;相比之下,在CrFeCoNiPd合金中,钯原子的原子尺寸和电负性与其他元素明显不同,其均质性大大降低。映射原子级元素分布为理解化学结构提供了机会,从而为调整组成和原子构型以获得出色的机械性能提供了基础(点击阅读);
【4】量子计算的成功依赖于纠缠大规模系统的能力。多方纠缠态对于量子信息科学中的众多应用至关重要。然而,在完全可控和可扩展的量子平台上生成和验证多方纠缠仍然是一个突出的挑战。2019年8月9日,浙江大学王浩华,中国科学院物理研究所范桁及郑东宁共同通讯在Science 在线发表题为“Generation of multicomponent atomic Schr?dinger cat states of up to 20 qubits”的研究论文,该研究在量子处理器上报告了18-qubit Greenberger-Horne-Zeilinger(GHZ)状态和多组分原子薛定谔猫状态的确定性生成,其具有20个超导量子位。该研究在固态平台上的方法不仅应该激发对探索量子多体系统基本物理学的兴趣,而且还能够开发实际量子计量学和量子信息处理中的应用(点击阅读);【5】2019年4月12日,中科院上海药物所徐华强,王明伟,浙江大学张岩及匹兹堡大学医学院Jean-Pierre Vilardaga共同通讯在Science发表题为“Structure and dynamics of the active human parathyroid hormone receptor-1”的研究论文,该研究报告了人类PTH1R与长效PTH类似物和刺激性G蛋白结合的冷冻电子显微镜结构。 结合的肽采用延伸的螺旋,其氨基末端深入插入受体跨膜结构域(TMD),导致跨膜螺旋6的羧基末端部分解旋并在该螺旋中间诱导尖锐的扭结以允许受体 与G蛋白结合。与单个TMD结构状态相反,细胞外结构域采用多种构象。这些结果提供了对PTH结合和受体激活的结构基础和动力学的见解。总而言之,该结构模型有助于解释甲状旁腺激素如何与其受体相互作用以及受体激活的分子基础(点击阅读);
【6】最近提出遗传补偿反应(GCR)作为基因敲除和基因敲除之间表型差异的可能解释;然而,GCR的潜在分子机制仍然没有被描述。浙江大学彭金荣及陈军在Nature在线发表题为“PTC-bearing mRNA elicits a genetic compensation response via Upf3a and COMPASS components”的研究论文,该研究使用capn3a和nid1a基因的斑马鱼敲除和敲除模型,显示带有过早终止密码子(PTC)的mRNA迅速触发涉及Upf3a和COMPASS复合物组分的GCR。与具有小肝脏的capn3a敲低胚胎和具有短体长的nid1a敲低胚胎不同,capn3a-null和nid1a-null突变体看起来正常,这些表型差异归因于同一家族中其他基因的上调。这些发现为GCR提供了潜在的机制基础,并且可能有助于开发治疗策略,通过在突变基因中产生PTC或引入含有PTC的转基因来触发GCR来治疗与遗传病相关的错义突变(点击阅读);
【7】2019年2月28日,哈佛医学院吴皓,James J. Chou及浙江大学陈枢青共同通讯在Cell发表题为“Higher-Order Clustering of the Transmembrane Anchor of DR5 Drives Signaling”的研究论文,该论文报告了一个意外的发现,对于死亡受体5(DR5),肿瘤坏死因子受体超家族中的受体,受体中单独的跨膜螺旋(TMH)直接组装高阶结构以驱动信号传导,并且这种结构未被束缚的胞外域抑制。该研究提供了新的机会和独特的观点来调节这些受体的信号转导,这些数据可用于疾病治疗,包括癌症免疫疗法(点击阅读);
【8】2019年1月18日,南京大学汪民怀,浙江大学俞绍才等人在Science上发表了题为“Aerosol-driven droplet concentrations dominate coverage andwater of oceanic low level clouds”的文章,该研究表明气溶胶驱动的液滴浓度是主导海洋低层云的覆盖和水的重要因素(点击阅读);【9】2019年1月9日,浙江大学陈红胜,新加坡南洋理工大学张柏乐及Gao Zheng共同通讯在Nature在线发表题为“Realization of a three-dimensional photonic topological insulator”的研究论文,该论文实验证明了一种具有极宽(超过25%带宽)3D拓扑带隙的3D光子拓扑绝缘体。使用直接场测量,研究人员绘制出有间隙的体带结构和光子表面态的狄拉克样色散,并展示沿非平面表面的稳健光子传播。该工作将3D拓扑绝缘体系列从费米子扩展到玻色子,并为三维几何中的拓扑光子腔,电路和激光器的应用铺平了道路(点击阅读);
1.人类阳离子-氯化物共转运蛋白KCC1的低温EM结构

人类溶质载体12(SLC12)基因家族编码阳离子-氯化物共转运蛋白(CCC),该转运蛋白介导Na +和/或K +与Cl-跨质膜的耦合电子中性共转。根据其转运特性和氨基酸序列定义,CCC可分为几个分支,包括两个Na-K-2Cl协同转运蛋白(NKCC1和NKCC2),一个Na-Cl协同转运蛋白(NCC)和四个K-Cl协同转运蛋白(KCC1- KCC4)。
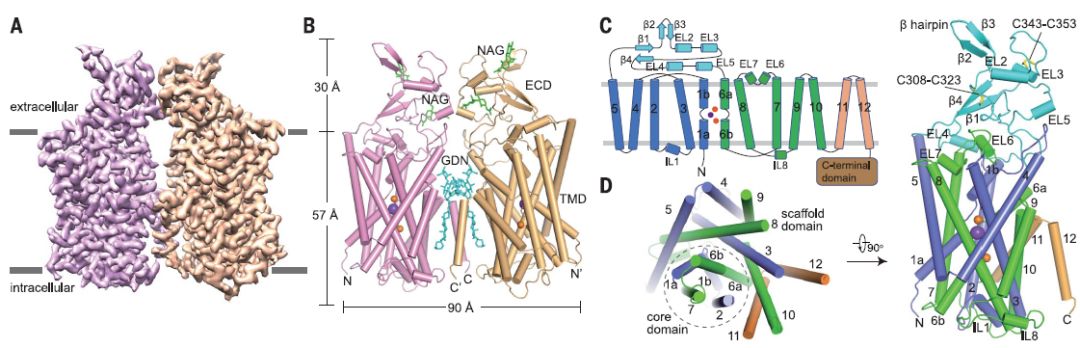
KCl和GDN中确定的人类KCC1的总体结构
CCC在许多组织和器官中表达,尤其是在肾脏和神经系统中,并在多种生理过程中发挥重要作用。当细胞暴露于高渗或低渗环境时,NKCC和KCC会通过跨质膜运输离子来减少细胞的渗透压差异,从而参与细胞体积调节。在肾脏中,NCC和NKCC2分别是远端回旋小管和Henle的厚上升环中NaCl重吸收的主要载体。在中枢神经元中,NKCC1和KCC2共同设定细胞内Cl-的浓度,从而调节γ-氨基丁酸(GABA)-和甘氨酸介导的突触抑制。
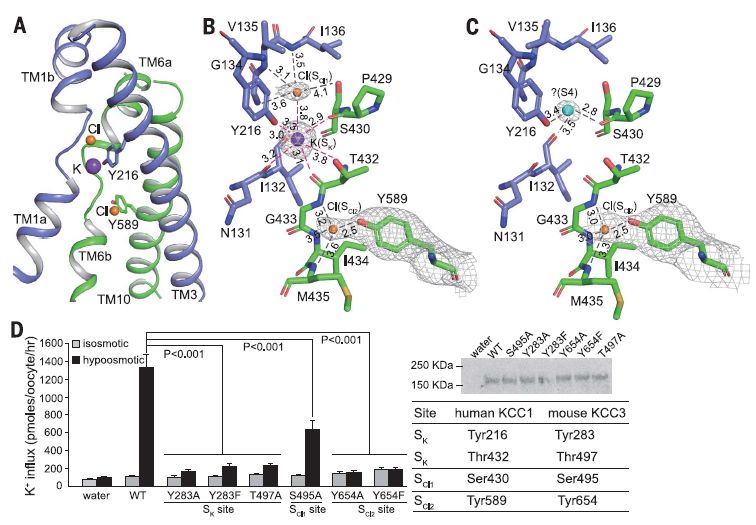
KCC1中的离子结合位点
考虑到它们在离子运输中的重要功能,CCC的突变会导致多种人类疾病也就不足为奇了。例如,KCC2中的五个单个氨基酸取代可导致人类癫痫。因此,CCC是重要的药物靶标。利尿剂和噻嗪分别通过抑制NKCC2和NCC用作降压药。最近,已经做出努力来开发用于治疗癫痫病的KCC2特异性激动剂和NKCC1特异性拮抗剂。通过调节GABA能抑制,靶向KCC2或NKCC1的药物也可能有效治疗其他疾病。例如,KCC2激动剂CLP290促进了脊髓损伤后的功能恢复。

团队成员郭江涛、叶升和刘斯在讨论课题
尽管已广泛研究了不同CCC成员的生理学,生物化学和药理学,但由于缺乏高分辨率结构,我们对其离子共转运机制的了解受到了阻碍。尽管DrNKCC1结构揭示了钠驱动CCC的结构和离子结合位点,但钾驱动KCC的结构和离子共转运机理仍然未知。
该研究介绍了在2.9至3.5埃分辨率下,在氯化钾或氯化钠中人氯化钾共转运蛋白KCC1的冷冻电子显微镜(cryo-EM)结构。KCC1以二聚体形式存在,胞外和跨膜结构域均参与二聚化。 结构和功能分析以及计算研究表明,KCC1中有一个钾位和两个氯化物位,这些都是离子迁移活性所必需的。 KCC1采用向内构象,并封闭了细胞外门。KCC1结构使我们能够对KCC中潜在的离子传输机制进行建模,并为药物设计提供蓝图。
KCC1的结构和功能表征使我们能够获得K-Cl共运模型。KCC1结构为解释其他KCC(例如KCC2和KCC3)的疾病相关突变提供了框架,这些突变会引起人类神经系统的遗传疾病。 人类KCC1与斑马鱼NKCC1的结构研究将促进针对人类CCC的药物开发和疾病治疗。
参考消息:
https://science.sciencemag.org/content/366/6464/505
2.细菌感测需要NOD1和NOD2的棕榈酰化

胞质模式识别受体(PRRs)核苷酸低聚结构域1(NOD1)和NOD2在宿主防御和存活中起关键作用,主要是通过赋予对胞质细菌肽聚糖[γ-D-谷氨酰基-间-二氨基庚二酸(iE-DAP)]以及在感染过程中细菌散发出的鼠疫二肽(MDP)的响应。NOD1 / 2功能失调会导致严重的免疫和炎症性疾病,例如克罗恩病(CD)和布劳综合征。 NOD2变异(3020insC移码突变,导致截短的NOD2蛋白)与CD的发病机理有关。尽管可溶于胞浆,但NOD1 / 2与质膜(PM)和内体区室结合,用于监测细菌细胞壁成分,并促进核因子κB(NF-κB)的活化和促分裂原的活化蛋白激酶MAPK)通过RIP2激酶从内体膜传递信号的途径。
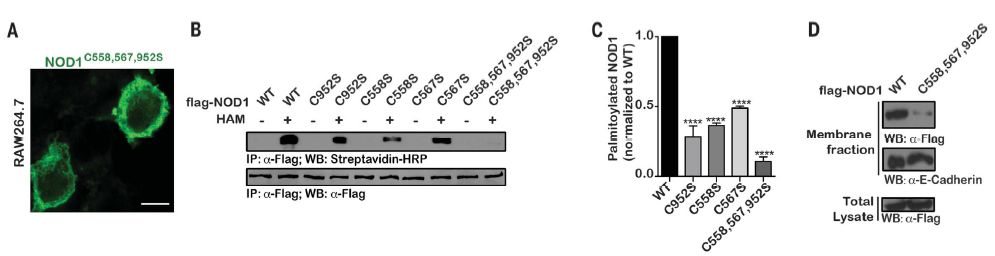
NOD1中的多个半胱氨酸残基可被酰化
虽然完全有能力结合MDP,但在NOD23020insC变体中未观察到膜缔合,这表明NOD1 / 2的膜定位对于其功能至关重要。缺乏可识别的膜靶向结构域,NOD1 / 2被建议通过细胞骨架成分或膜结合蛋白间接锚定于膜,或通过内体蛋白如SLC15A3锚定于内体。但是,这些模型不能完全解释由于细菌入侵,NODs从质膜到内体区室的快速重新分布。
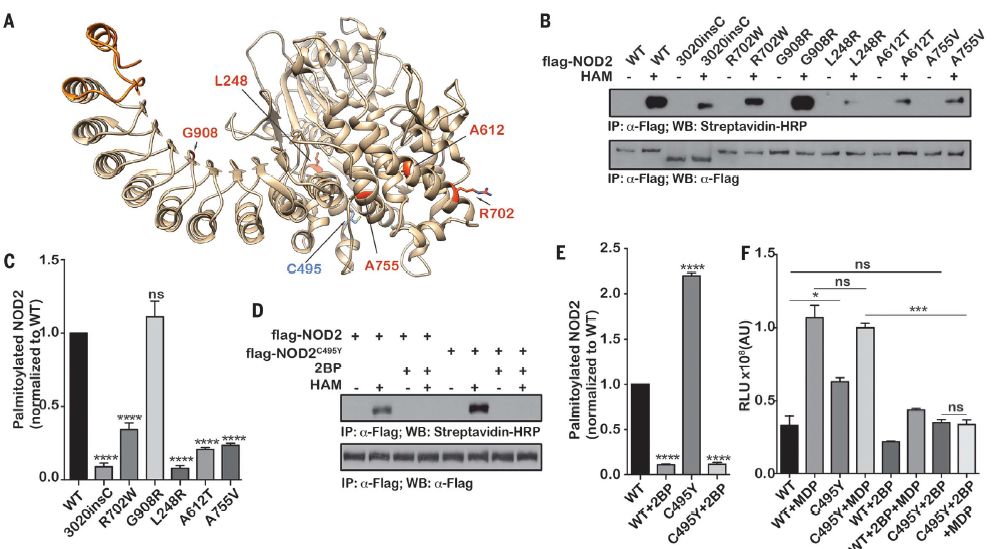
NOD2编码变体表现出异常的S-棕榈酸酯化
可溶性蛋白可以通过脂化作用靶向细胞膜结构。与法呢基化或香叶基化不同,棕榈酰化不需要目标蛋白中的共有基序,并且广泛参与蛋白质定位,运输和稳定性的调节。含有ZDHHC结构域的蛋白质酰基转移酶(PAT)催化S-棕榈酰化,在免疫应答中也起着重要作用。
该研究发现膜募集和免疫信号传导需要NOD1 / 2 S-棕榈酰化。ZDHHC5被确定为负责此关键的翻译后修饰的棕榈酰转移酶,并且发现NOD2中的几种与疾病相关的突变与缺陷性S-棕榈酰化有关。因此,ZDHHC5介导的NOD1 / 2的S-棕榈酰化对于它们对肽聚糖的应答和进行有效免疫应答的能力至关重要。

Dante Neculai指导团队成员分析结果
该研究发现将促进对NOD引起的炎性疾病的理解,并最终对包括NOD依赖性自身免疫性疾病和慢性细菌感染进行治疗。
参考消息:
https://science.sciencemag.org/content/366/6464/460


